(通讯员 朱潇潇)近日,国际著名综合性期刊Proceedings of the National Academy of Sciences,PNAS(美国科学院院刊,影响因子:12.78)发表了华中师范大学生命科学学院罗金红教授课题组和武汉大学测绘学院李星星教授课题组的合作研究成果,论文题目为“Sensory Error Drives Fine Motor Adjustment”(https://doi.org/10.1073/pnas.2201275119)。华中师范大学研究生汪慧敏、武汉大学研究生周宇轩为论文的共同第一作者,武汉大学李星星教授和华中师范大学罗金红教授为论文的共同通讯作者。
提到蝙蝠,大家首先想到的关键词可能是“暗夜行者”。与猫头鹰独特的夜视能力不同,蝙蝠主要依靠回声定位(也称生物声呐)称霸夜空。据调查,全世界有超过1400种蝙蝠,其中回声定位蝙蝠约1200种。回声定位蝙蝠依靠声呐进行捕食、躲避障碍物等活动。

回声定位蝙蝠利用声呐进行捕食1
蝙蝠的回声定位行为很好地说明了听觉反馈-发声控制的重要性。几乎所有的回声定位蝙蝠都展现出“快而准”的适应性发声调节行为1,2。其中,分布在旧大陆的菊头蝠和蹄蝠展现出极高的声波频率控制精准度,约是专业歌唱家音调控制精准度的5倍3–5。
蝙蝠作为哺乳动物的一员,与人类共享基本的大脑解剖结构,因其具备罕见的回声定位能力而显得格外特殊。此外,绝大多数回声定位蝙蝠也通过与人和其他哺乳动物相似的喉部来发声。因此,一直以来回声定位蝙蝠都是研究发声控制和听觉的主要哺乳动物模型之一。
近期,罗金红教授课题组以大蹄蝠(Hipposideros armiger)为实验对象,利用听觉反馈干扰行为范式解析了大蹄蝠发声频率调节的计算机制。实验人员基于FPGA(可编程逻辑门阵列)技术首次搭建了声信号输入–输出时延低于千分之一秒的实时变频听觉反馈干扰平台,从而可以在线调节大蹄蝠接收到的听觉反馈信息。通过创造听觉感觉误差,研究人员发现大蹄蝠呈现出了快速的频率补偿行为——发声频率改变方向与干扰频率(感觉误差)方向相反。
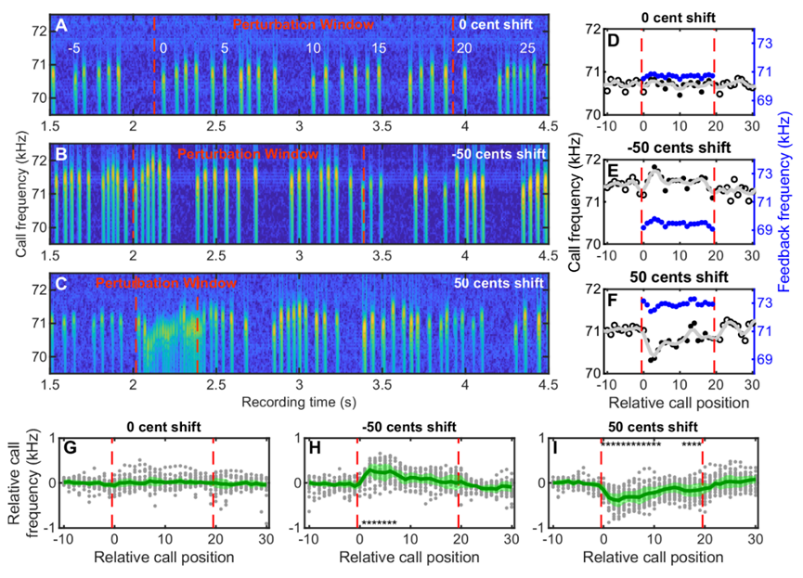
大蹄蝠发声频率改变方向与频率干扰方向相反,呈现出频率补偿行为
该研究进一步发现,干扰频率(感觉误差)的大小影响大蹄蝠频率补偿的大小,但是两者并不是简单的线性关系。其中在正向50音分的频移下,大蹄蝠的频率补偿行为最为明显。这可能是因为正向50音分对应的飞行速度与大蹄蝠在自然状态下的飞行速度最接近,蝙蝠对此实验条件感觉更真实。
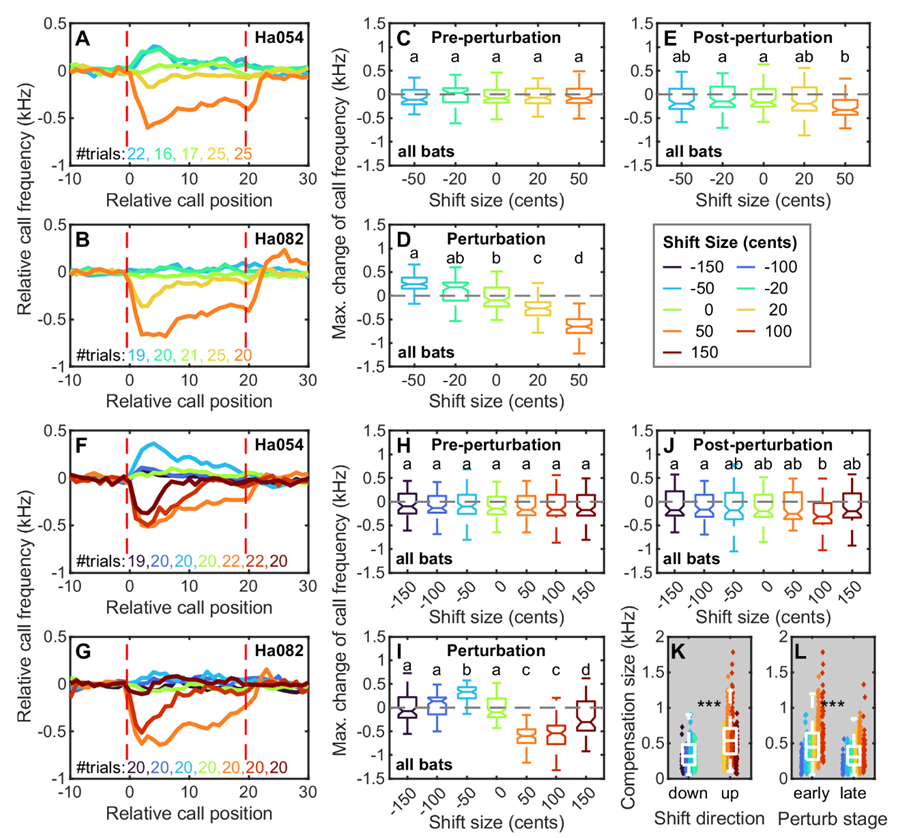
大蹄蝠在不同干扰频率下的频率补偿
目前关于回声定位蝙蝠发声频率调节的假说主要包括多普勒频移补偿(Doppler shift compensation)、干扰规避反应(Jamming avoidance response)和注意反应假说(Attention reaction hypothesis),但是这些现有的假说均不能完整地解释本实验的结果。“难道蝙蝠回定位声波频率的控制并不是基于回声定位蝙蝠特有的机制?”随后,研究人员将思路投向了人类言语的频率调节假说——感觉误差校准假说(Sensory error correction)6,7。人类言语的频率调节机制研究相对透彻,已经建立了高质量的数据计算模型。为了验证大蹄蝠频率调节机制是否与人类言语的感觉误差校准假说类似,课题组与武汉大学测绘学院的李星星教授团队一拍即合,开始了深入的交流与合作。经过不懈努力,成功将人类言语频率调节的状态反馈控制(State feedback control)模型应用到了大蹄蝠回声定位声波频率调节上。在大蹄蝠上构建的状态反馈控制模型不仅具有极高的精度(总体误差仅为61 Hz,约为大蹄蝠自然发声频率变异的四分之一),而且能解释为什么同一只蝙蝠在不同听觉反馈干扰条件下呈现出不同的发声频率调节行为,也能够解释同一条件下不同个体之间的显著行为差异。
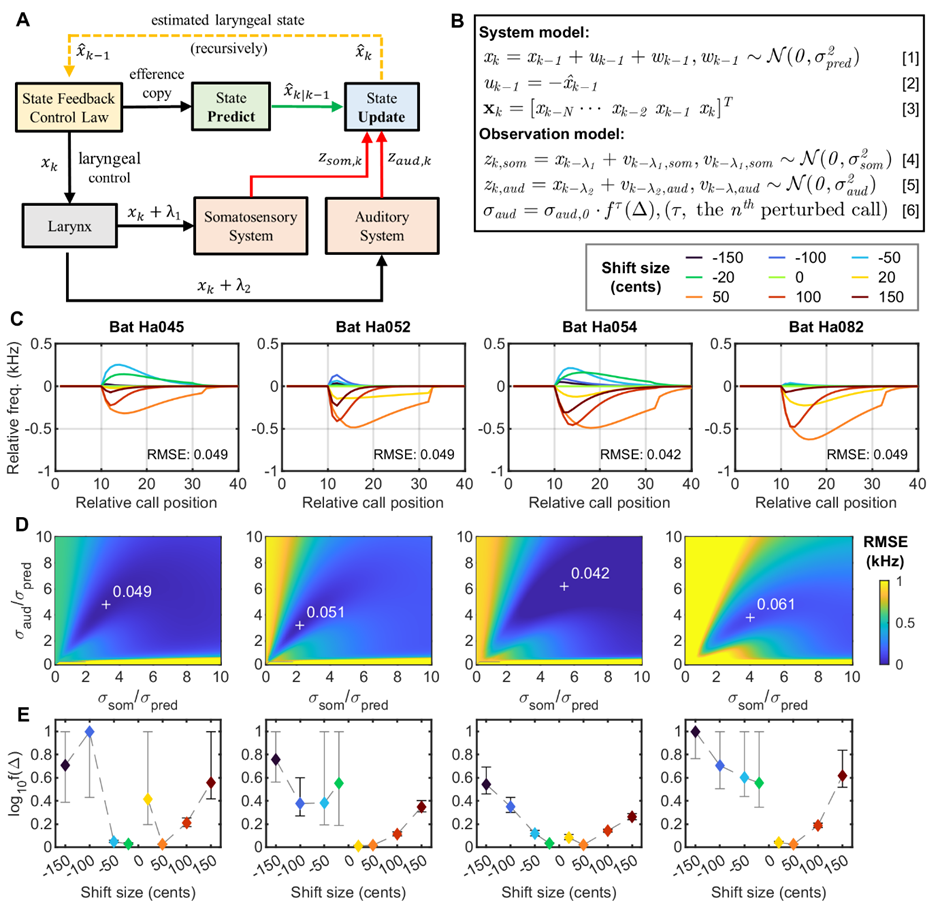
大蹄蝠发声频率调整的计算模型
该研究不仅揭示了感觉误差是驱动回声定位蝙蝠动态发声调节的基本机制,也说明尽管蝙蝠回声定位和人类言语的功能完全不同,但是它们在听觉反馈-发声控制方面依赖共同的计算原理。
该研究得到了国家自然科学基金委面上项目、国际人类前沿科学“职业发展奖”等项目的资助。
参考文献:
1. Luo, J., and Moss, C.F. (2017). Echolocating bats rely on audiovocal feedback to adapt sonar signal design. Proc. Natl. Acad. Sci. U.S.A. 114, 10978–10983.
2. Luo, J., Kothari, N.B., and Moss, C.F. (2017). Sensorimotor integration on a rapid time scale. Proc. Natl. Acad. Sci. U.S.A. 114, 6605–6610.
3. Schnitzler, H.U., and Denzinger, A. (2011). Auditory fovea and Doppler shift compensation: adaptations for flutter detection in echolocating bats using CF-FM signals. J. Comp. Physiol. A. 197, 541–559.
4. Zhang, Y., Lin, A., Ding, J., Yang, X., Jiang, T., Liu, Y., and Feng, J. (2019). Performance of Doppler shift compensation in bats varies with species rather than with environmental clutter. Anim. Behav. 158, 109–120.
5. Kishon-Rabin, L., Amir, O., Vexler, Y., and Zaltz, Y. (2001). Pitch discrimination: are professional musicians better than non-musicians? J Basic Clin Physiol Pharmacol 12, 125–143.
6. Liu, H., and Larson, C.R. (2007). Effects of perturbation magnitude and voice F0 level on the pitch-shift reflex. The Journal of the Acoustical Society of America 122, 3671–3677.
7. Parrell Benjamin and Houde John (2019). Modeling the role of sensory feedback in speech motor control and learning. Journal of Speech, Language, and Hearing Research 62, 2963–2985.
8. Wang, H., Zhou, Y., Li, H., Moss, C.F., Li, X., and Luo, J. (2022). Sensory error drives fine motor adjustment. Proc Natl Acad Sci USA 119, e2201275119.
(审读人 谢波)
©版权所有 :华中师范大学科技处


